Home » 2019
Yearly Archives: 2019
Marine Geology field trip to Nijar and Sorbas
As the last year, students of the third course of Marine sciences of the Alicante University visit the Sorbas basin, Cabo de Gata, Hoyazo de Nijar, and Cerro del Espíritu Santo of Vera. The visit took place on 7 and 8 of November of 2019.
The visit was organized by Antonio Estévez, Santiago Moliner, and José Enrique Tent-Manclús.

Student group before entering the Sorbas Cave.
Alicante Messinian Group LOGO

The logo was made by Jesús M. Soria based in an eroded mediterranean margin resembling the M of Messinian.
The “Paleoenviromental changes” logo is for the Research Group of the Alicante University.
The Alicante Messinian Group was granted by the Ministerio de Ciencia e Innovación Research Project CGL 2007 -6583 .
Neogene Brachiopods and Betic Corridors
Diego García-Ramos
University of Vienna, Institut for Paläontologie
1.Introduction
Brachiopods, a name that derives from Ancient Greek words βραχίων (“arm”) and πούς (“foot”), make up a phylum of tripoblastic, coelomate, marine invertebrates that belong, along with bryozoans and phoronideans, to the superphylum of the so-called lophophorates. Their body plan consists of two valves displaying bilateral symmetry, and are generally inequivalve. Most brachiopods are sessile, stenohaline suspension feeders, and their fossil record spans from Lower Cambrian times to Recent. Today’s brachiopod diversity shows that they constitute a relic group of animals when compared with their paleobiodiversity during past geological time, especially in the Paleozoic, throughout which brachiopods were extremely diverse and abundant. Nowadays there are some 391 extant species of which around 5 % may be synonyms (Emig et al., 2013). Extant brachiopods are distributed from the intertidal zone to abyssal depths, but they occur more frequently in the inner and outer shelf, and the epibathyal zone.
- Brachiopods as paleocological and paleobiogeographical proxies
Since brachiopods are overall facies dependent organisms (Ager, 1965), together with the fact that many of their species display a long-ranging vertical distribution, their use as guide-fossils is limited in comparison with other groups like ammonoids and calcareous micro- and nannoplankton, despite the fact that many Paleozoic brachiopods are important biostratigraphical markers. In any case, the precise feature that they are facies-linked animals make brachiopods considerably suitable for paleoecological and paleogeographical studies. In particular, Ager (1967) outlined a number of reasons why fossil brachiopods may be of great use in paleoecological studies: 1) their great abundance throughout the Phanerozoic; 2) their occurrence in diverse sedimentary facies; 3) their susceptibility to environmental differences and perturbation; 4) the fact that their skeletal traits record physiological differences which are related to their life habit; and 5) the survival, with a relatively adequate diversity, of modern representatives which may aid in actuopaleontological analyses.
As for their use in paleobiogeography, Smirnova (2012) stressed the fact that brachiopods, most of which were/are fundamentally shelf dwellers, are good proxies to estimate the proximity of the coastline, and enable to contour marine basins accurately. Other than that, she also pointed out that brachiopods are sensitive to climatic factors, and the taxonomic composition of brachiopods has been proven to be a good tool in differentiating paleobiogeographical realms.
- Overview of Tertiary brachiopods from the Betic Cordillera
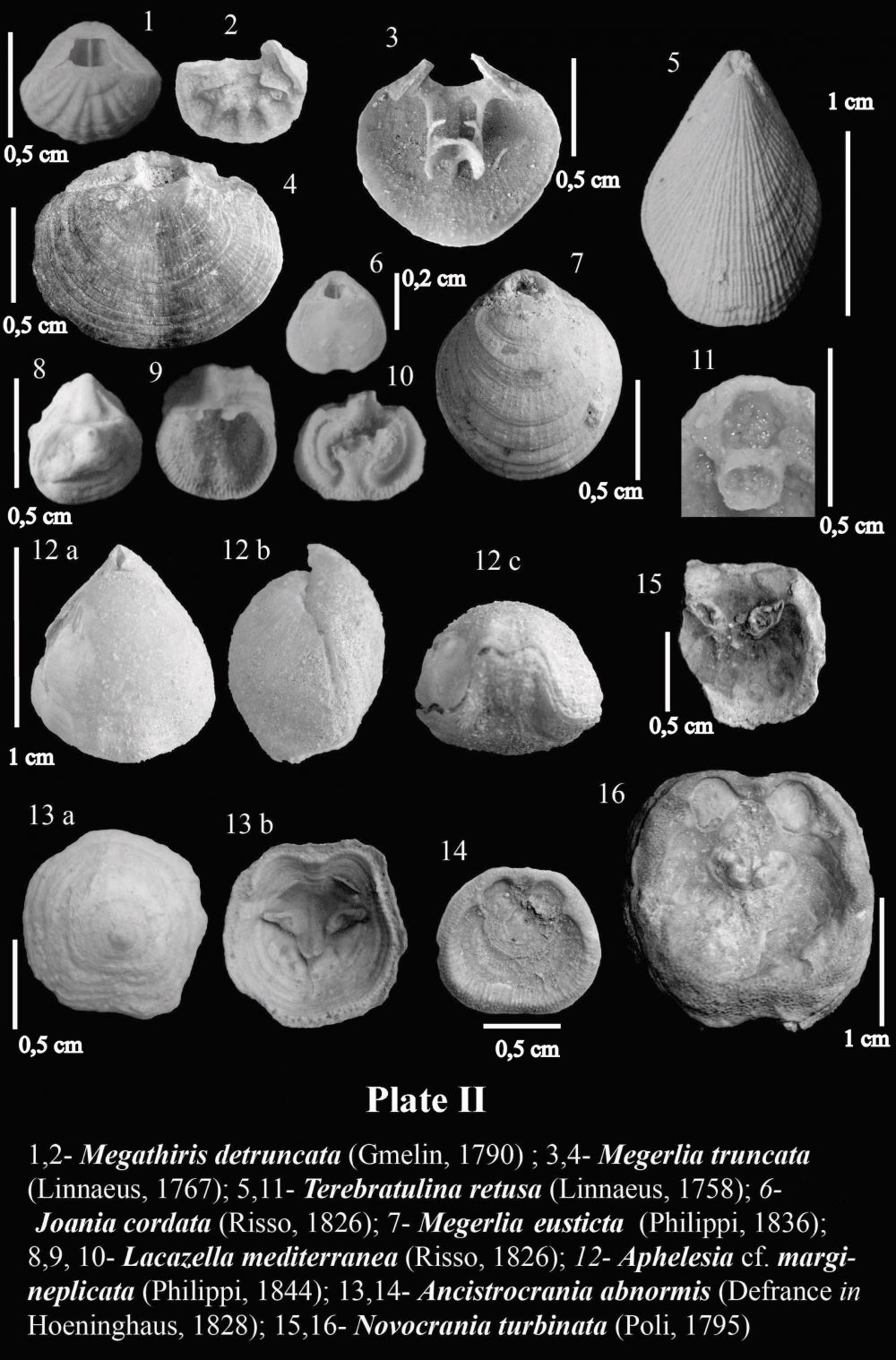
Brachiopods in the Tertiary record of the Betic Cordillera are relatively poorly known. There are sparse citations in old publications such as that of Davidson (1864), who mentioned the occurrence of Megerlia truncata(Linnaeus, 1767) in the Miocene of Gibraltar, or Terebratula sinuosa (Brocchi, 1814) in Miocene rocks from Córdoba (Davidson, 1870) (later known to be Terebratula maugerii Boni, 1933). Kilian in Fouqué (1884) cited a number of species of Terebratula, besides the rhynchonellid and thecideid brachiopods Aphelesia bipartita(Brocchi, 1814) and Lacazella mediterranea (Risso, 1826), from the Neogene record of the Granada Basin.
A general glance at Cainozoic (but mainly Neogene) brachiopod paleobiodiversity in the Betic Cordillera shows that the genus Terebratula Müller, 1776 prevails by far, making up more than 80% of the record of brachiopods during this era (Toscano et al., 2010). In rocks of Eocene age, terebratulids attributable to Carneithyris hilarionis (Davidson, 1870) are known to occur rarely in the province of Alicante (Sulser et al., 2010), and there is unpublished record of Upper Oligocene to Lower Miocene small gryphine brachiopods, along with Terebratulina gr. tenuistriata (Leymerie, 1846), in the province of Murcia.
Two main types of brachiopod assemblages may be recognized in Neogene deposits of the eastern Betic Cordillera: a relatively shallow water Terebratula–Aphelesia assemblage associated to fine-grained bottoms, forming Terebratulapavements or clumps, with sparse, solitary occurrence of Aphelesia, and a bathyal assemblage where small gryphine-like terebratulids and Ceramisia meneghiniana (Seguenza, 1865) are distinctive components, found in deep-sea hard-substrates and associated to bamboo-coral bioherms (see Barrier et al., 1991). Other taxa such as Megerlia truncata, Terebratulina retusa (Linnaeus, 1758) and Lacazella mediterranea display an euribathic distribution and may occur in both assemblages as long as there is suitable substrate. The micromorphic brachiopod Megathiris detruncata (Gmelin, 1790) and the inarticulate Ancistrocrania abnormis(Defrance in Hoeninghaus, 1828) have been found in infralittoral to circalittoral hard-substrates consisting either of shelly-grounds or rocky-grounds. Joania cordata (Risso, 1826) and Novocrania turbinata (Poli, 1795) have exclusively been found in hard-substrates corresponding to a circalittoral volcanic swell.
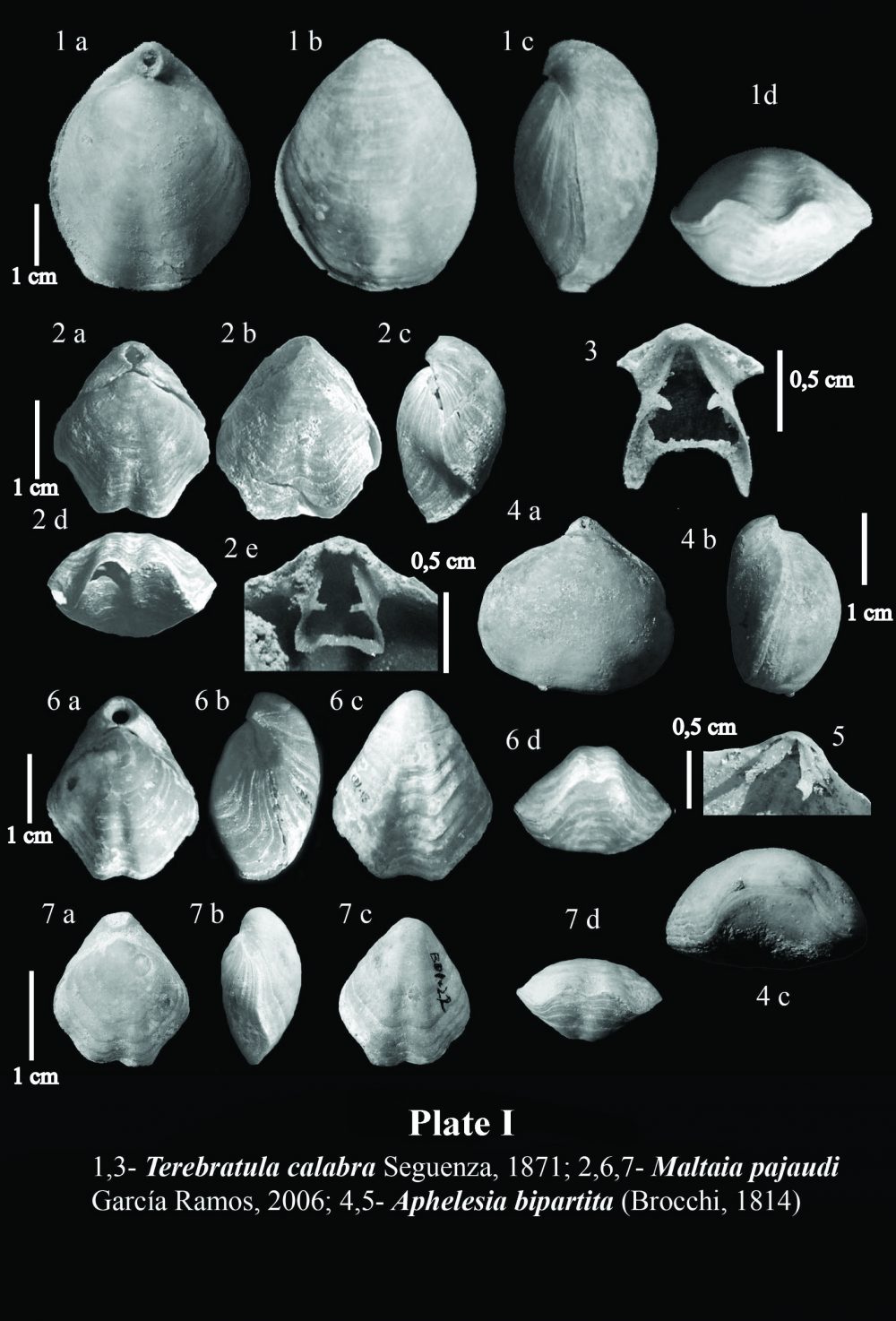
In sediments of Late Neogene age deposited the intramountaneous basins of the Betic Cordillera, the recurrent assemblage of species of the genera Terebratula -or its close relative Maltaia Cooper, 1983-and Aphelesia Cooper, 1959, has been already pointed out elsewhere by, for instance, Pedley (1976) and Gaetani (1986). During the Tortonian, the species Aphelesia margineplicata(Philippi, 1844) commonly co-occurred in silty to fine-grained sandy lithofacies in association with Terebratula maugeriior Terebratula pseudoscillae Sacco, 1902 (see also Calzada, 1978). In addition, it has also been found in the same outcrops together with Miocene representatives of Terebratula calabra Seguenza, 1871, or Maltaia aff. costae (Seguenza, 1871), in Late Tortonian to Early Messinian aged deposits, mainly interpretable as of infralittoral to circalittoral paleoenvironments.
The onset of the Messinian Salinity Crisis exerted a profound impact on practically the whole marine paleobiota of the Mediterranean at the end of Messinian times, and brachiopods were not an exception, but they were among the victims of this vast extinction. The restoration of normal marine conditions that followed the opening of the Strait of Gibraltar, leading to the Early Pliocene re-flooding, implied a re-colonization by brachiopods, ever since Terebratula and Aphelesia had survived in Atlantic waters (see, for example, Dollfus & Cotter, 1909 and Toscano et al., 2010). The taxonomic composition of Pliocene Terebratula–Aphelesiaassemblages of the Betic Cordillera marked a faunal turnover when compared to those prevailing during the Miocene. The species Aphelesia bipartita replaced A. margineplicata –which presence became marginal- as the dominating partner of Terebratula (e.g. Pajaud, 1976). Likewise, T.pseudoscillae and the once abundant Terebratula maugerii= T. sinuosadied out. Their niche was taken over by Terebratula terebratula (Linnaeus, 1758), by Pliocene morphotypes of Terebratula calabra, and by Maltaia pajaudi García-Ramos, 2006.

4.Brachiopods and Betic Corridors
Reolid et al. (2012) have shown that the genus Terebratula might have likely thriven in the shelf margins of Atlantic-Mediterranean corridors by benefitting from warmer and more saline water currents flowing from the Mediterranean to the Atlantic. The occurrence of Terebratula in Betic intramountane basins that might perform as Atlantic-Mediterranean seaways can be traced as far back as to Early Miocene times (e.g. García-Ramos, 2006). Llarena (1934) mentioned the presence of Terebratula sp. and Terebratula grandis (probably T. pseudoscillae) in basins located within the Prebetic realm of the province of Albacete, and Darder Pericás (1945) likewise described outcrops yielding numerous specimens of Terebratula in Neogene basins of the Prebetic in the southern sector of the province of Valencia. All of these basins might be part of an interconnected web of Atlantic-Mediterranean seaways. Further studies are needed to solve the puzzle as to how the Atlantic-Mediterranean connections evolved in the Betic Cordillera as the Alborán Block was undergoing indentation against the Iberian paleomargin. The study of marine epibenthic macroinvertebrates, including brachiopods, may play a role in enhancing our knowledge of how all of this happened.
References:
Ager, D.V., 1965, The adaptation of brachiopods to different environments: Palaeogeography, Palaeoclimatology, Palaeoecology, 1, p. 143-172.
Ager, D.V., 1967, Brachiopod palaeoecology: Earth-Science Review, 2, p. 157-159.
Barrier, P., Zibrowius, H., Lozouet, P., Montenat, C., Ott d’Estevou, P., Serrano, F., Soudet, H. J., 1991, Une faune de fond dur du bathyal superieur dans le Miocene terminal des Cordilleres Betiques (Carboneras, SE Espagne): Mesogee (Bulletin du Museum d’Histoire Naturelle de Marseille), 51, p. 3-13.
Bertrand, M., Kilian, W., 1884, Études sur les terrains secondaires et tertiaires dans les provinces de Grenade et de Malaga, in : «Mission d’Andalousie » : études relatives au tremblement de terre du 25 décembre 1884 et à la constitution géologique du sol ébranlé par les secousses: Mém. Acad. Sci. Paris, 30 (2), p. 377–579.
Calzada B. S., 1978, Braquiópodos tortonienses de Murcia: Estudios geológicos, 34, p.351-358.
Davidson, T., 1864, Description of the Brachiopoda: in Adams S L. (ed.), Outline of the geology of the Maltese Islands. The Annals and Magazine of Natural History, Series 3, 14, p. 5-11.
Davidson, T., 1870, On Italian Tertiary Brachiopoda: Geological Magazine, London, 7 (8), p. 359-370, p. 399-408, p. 460-466.
Darder Pericás, B., 1945, Estudio geológico del sur de la Provincia de Valencia y norte de la de Alicante. Boletín Geológico y Minero de España, 57: p. 59-837.
Dollfus, G.F. & Cotter, J.C.B, 1909, Le Pliocène au Nord du Tage (Plaisancien).Brachiopoda. Commission du service géologique du Portugal, p. 87-90.
Emig, C. C., Bitner, M.A., Álvarez, F., 2013, Phylum Brachiopoda. In: Zhang, Z.-Q. (Ed.) Animal Biodiversity: An Outline of Higher-level Classification and Survey of Taxonomic Richness (Addenda 2013). Zootaxa, 3703, 1–82.
Gaetani, M., 1986, Brachiopod paleocommunities from the Plio-Pleistocene of Calabria and Sicilia (Italy). In: Biostratigraphie du Paléozoïque 4. Les brachiopodes fossils et actueles. P.R. Racheboeuf & Emig, C.C. (Ed.). p. 281-288.
García Ramos, D.A. 2006. Nota sobre Terebratulinae del Terciario de Europa y su relación con los representantes neógenos del sureste español. Boletín de la Asociación Cultural Paleontológica Murciana 5: 23-83.
Llarena, J.G. de, 1934, Observaciones sobre la geología y fisiografía de los alrededores de Hellín. Boletín de la Real Sociedad Española de Historia Natural, p.213-231.
Pajaud, D., 1976, Les Brachiopodes du Pliocène I de la Sierra de Santa Pola (sud d’Alicante, Espagne): Terebratula terebratula (Linné, 1758) et Phapsirhynchia sanctapaulensis nov. gen., nov. sp.: Annales de la Société géologique du Nord, 96, p. 99-106.
Pedley H. M., 1976, A palaeoecological study of the Upper Coralline Limestone, Terebratula-Aphelesia Bed (Miocene, Malta) based on bryozoan growth-form studies and brachiopod distributions: Palaeogeography Palaeoclimatology Palaeoecology, 20, p. 209-234.
Smirnova, T.N., 2012, Early Cretaceous Rhynchonellids of Dagestan: System, Morphology, Stratigraphic and Paleobiogeographic Significance. Paleontological Journal, v.46, nº 11, p. 1197-1296.
Reolid, M., García-García, F., Tomasovych, A. and Soria, J.M., 2012, Thick brachiopod shell concentrations from prodelta and siliciclastic ramp in a Tortonian Atlantic–Mediterranean strait (Miocene, Guadix Basin, southern Spain). Facies 58: 549-571.
Sulser, H., García-Ramos, D., Küsteiner, P., Menkveld-Feller, U., 2010, Taxonomy and palaeoecology of brachiopods from the South-Helvetic zone of the Fäneren region (Lutetian, Eocene, NE Switzerland). Swiss Journal of Geosciences, 103: p. 257–272.
Toscano-Grande, A., García-Ramos, D., Ruiz-Muñoz, F., González-Regalado, M.L., Abad, M., Civis-Llovera, J., González-Delgado, J.A., Rico-García, A., Martínez-Chacón, M.L., García, E.X. and Pendón-Martín, J.G. 2010. Braquiópodos neógenos del suroeste de la depresión del Guadalquivir (sur de España). Revista Mexicana de Ciencias Geológicas 27: 254-263.
This post is an extended abstract of the work by Diego García-Ramos (2006): Nota sobre terebatulinae del Terciario de Europa y su relación con los representantes neógenos del sureste español. Boletín de la Asociación Cultural Paleontológica Murciana. 5, pp. 23-83
Villafranqueza corals in the 34th IAS Meeting of Sedimentology (Rome)
Entobia is a well-known ichnofacies typical for nearshore environments, differentiated from the Gnatichnus ichnofacies as being related to more stable conditions in high-energy settings and longer times of exposure of the bottoms. Surroundings of the city of Alicante (Villafranqueza section, SE Spain), boring structures related to the Entobia ichnofacies were observed on an Eocene rockground colonized after a Miocene marine ingression. Reefrelated facies are present as coral colonies developed after the transgression and show evidence of bioerosive activity. The relationship existing between the growing coral colony and boring organisms is addressed in order to elucidate ecological interactions and palaeoenvironmental conditions. The studied section is characterized by a very evident angular unconformity dividing the underlying Eocene sandy turbiditic beds from the overlying Miocene coastal limestones. The Entobia ichnofacies is developed on a variety of substrates, represented by the Eocene rockground, resedimented lithoclasts, different bioclasts (mainly thick shelled bivalves and gastropods) and coral bioconstructions. Boring structures are mainly represented by Gastrochaenolites, Entobia (usually preserved as internal mould) and, more rarely, Trypanites. Differences in the relative abundance of the three ichnotaxa were observed according to the substrate: i) on the rockground, Gastrochaenolites and Trypanites are the most abundant ichnotaxa, being Gastrochaenolites represented by different ichnospecies; ii) the three ichnotaxa are present in litho- and bio-clasts; iii) in corals, almost exclusively Gastrochaenolites torpedo is found. On the rockground, deep penetrating borings as Gastrochaenolites are differently preserved, being present complete forms together with partially eroded ones on the same surface. This would suggest that colonization occurred over long times, due to prolonged exposure of the bottoms. Together with possible ecological restrictions, a preservation bias induced by such a prolonged exposure must be considered in order to justify the relative abundance of the deep structures typical for the Entobia ichnofacies with respect to the shallow-tiered ones. Where Gastrochaenolites is associated to corals, bivalves were found inside the bioerosion structures being mainly represented by Lithophaga sp.. In this sense, a bioerosive activity specialism is detected, since Gastrochaenolites is extremely more abundant in the bioconstructions produced by the coral Tarbellastrea than in those where other types of corals are present (e.g. Porites and Siderastraea), thus suggesting some kind of ecological interplay. Moreover, the study of the geometrical and morphological relationship between Tarbellastrea and Gastrochaenolites shows that, in part of the observed specimens, the coral was growing all around the lithophagid borehole. This would suggest that corals and bivalves developed at the same time and that the shelly colonization and the progress of the host patch reef did not occur in different successive phases.
Cite as: Giannetti, A., Falces-Delgado, S., and Baeza-Carratalá, J. F. (2019): Interaction of boring organisms with corals. Entobia ichnofacies development in a transgressive nearshore scenario. 34th IAS International Meeting of Sedimentology, Rome 10-13 September 2019. “Sedimentology to face societal challenges on risk, resources and record of the past. Sesion 8.A-660.

The Villafranqueza unconformity in the logo of the Alicante University Departament of Earth Science and Environment.
Kuphus beds at the 34th IAS Meeting of Sedimentology
“Kuphus beds in an Oligocene carbonate platform (Sierra de la Argüeña, Southeastern Spain): palaeoecological context and relationship with shallowing-upward cycles” by Alice Giannetti and Santiago Falces-Delgado
Although the Oligocene carbonate platforms saw the development of Kuphus-dominated assemblages, few studies addressed the role of these bivalves in ancient shallow-water ecosystems. Kuphus is a genus of peculiar teredinid burrower bivalves that begun to be common in the Oligocene, reaching their maximum abundance in the Oligocene-Aquitanian time interval. Nowadays, they are represented only by the species K. polythalamia, present in the Western Pacific. Recent studies revealed that the physiology of K. polythalamiais based on symbiosis with chemoautotroph sulphur bacteria, this producing new interest about the role of Kuphus in ancient carbonate platforms. Kuphus siphons are embedded in concentric calcitic sheets forming long subcylindrical tubes, which accumulation produces thick beds with good preservation potential. In the Sierra de la Argüeña (Alicante province, Southeastern Spain), different types of Kuphus beds were recovered in nearshore carbonate platform deposits, preserved either as resedimented material or in situ assemblages. Five facies were distinguished: i. marly limestones rich in fragments of Kuphus tubes, corals, irregular echinoids, bivalves and gastropods; ii. miliolid-rich wackestones/packstones with small turriculate gastropods and the branched coral Stylophora, representing protected environments close to the coast; iii. small coral bioconstructions formed by laminar and domed colonies of Porites, Montipora, Actinastrea and different favids; iv. packstones rich in red algae and echinoid fragments with lepidocyclinids and miliolids, filling the space between coral bioconstructions; v. fine-grained greenish silty sands, locally associated to Microcodium. These facies are repeated throughout the section to form regressive cycles from the lagoon to coral patch reefs developed in the inner platform and then to nearshore areas, in proximity of the emerged coast. Beds with resedimented Kuphus tubes are found in the lowermost or in the uppermost part of the regressive sequences; in both situations, preferred orientation is absent. In the first case, they are made up of tube fragments mixed with other bioclasts and have been interpreted as perireefal accumulations. In the second case, the beds are made up almost exclusively of Kuphus tubes, show quite a high density of tube fragments and wide areal development. These have been interpreted as product of physical reworking, e.g. by storms. Several patches of tubes in life position vertically cross this bioclastic rudstones, suggesting that some bivalves survived the event and resumed the construction of the tube after the storm ceased. At the very top of the section, anterior ends are preserved in life position inside the fine-grained silty sands of facies v. This last level is of particular interest, since it represents the original undisturbed environment of the pioneer colonies. So far, accumulation of organic matter and plant remains similar to those present in the habitat of modern-day Kuphus were not observed in this bed, this suggesting the possibility that Oligocene Kuphus could have retained normal filter feeding mechanisms and were not obligate chemotrophic organisms. This would recommend prudence with the use of modern-day Kuphus polythalamia as an ecological analogue of fossil Kuphus and that further study is needed before considering the latter a proxy for poorly oxygenated environments.

Cite as: Giannetti, A., and Falces-Delgado, S. (2019): Interaction of boring organisms with corals. Entobia ichnofacies development in a transgressive nearshore scenario. 34th IAS International Meeting of Sedimentology, Rome 10-13 September 2019. “Sedimentology to face societal challenges on risk, resources and record of the past. Sesion 1.A-989.
Tethyan to Mediterranean Evolution
Message from the Guest Editor
Dear Colleagues,
Several geological modeling studies on the geodynamic and paleogeographic Alpine (Eo-Alpine during the Cretaceous–Paleogene; Neo-Alpine during the Oligocene– Miocene) evolution of the Mediterranean have been published without consensus in the last four decades. Therefore, a Special Issue dedicated to “Tethayan to Mediterranean evolution” seems justified. Studies that consider the geodynamic and/or paleogeographic evolution of the entire Mediterranean area or a part of this area will be welcome. The proposal can be focused on stratigraphic, sedimentological, petrographic, or tectonic data, or geophysical interpretations and reconstructions based on principles of Plate Tectonics using GPlates or similar software. The papers that are published in this Special Issue can contribute to clarifying and updating the state of our knowledge about this controversial theme.

Message from the Editorial Board
T h e Journal of Marine Science and Engineering (JMSE; ISSN 2077-1312) is an international peer-reviewed open access journal which provides an advanced forum for studies related to marine science and engineering. The journal aims to provide scholarly research on a range of topics, including ocean engineering, chemical oceanography, physical oceanography, marine biology and marine geosciences. We invite you to publish in our journal sharing your important research findings with the global ocean community.
Open Access: free for readers, with article processing charges (APC) paid by authors or their institutions.
High visibility: Indexed in the Science Citation Index Expanded in Web of Science, in Inspec (IET) and in Scopus.
CiteScore (2018 Scopus data): 1.76, which equals rank 30/92 in ‘Ocean Engineering’, rank 105/288 in ‘Civil and Structural Engineering’ and rank 77/203 in ‘Water Science and Technology’.
Guest Editor:
Prof. Dr. Manuel Martín-Martín
Earth sciences and environment
Department (University of Alicante). Campus Universitario del San Vicente del Raspeig, AP- 99, 03080 Alicante, Spain
manuel.martin.m3@gmail.com
Deadline for manuscript submissions:
10 March 2020
Sorbas and Nijar visit
Students of the third course of Marine sciences of the Alicante University visit the Sorbas basin, Cabo de Gata, Hojazo de Nijar, and Cerro del Espíritu Santo of Vera. The visit took place on 8 and 9 of November of 2018.
The visit was organized by José Enrique Tent-Manclús.

Photograph of students watching the sunset on the Monsúl beach by Carla Morales Pujalte.

Students watching the Monsul beach ignimbrite. Photography by Esther Ivars.
Members of the Alicante Messinian Group
The team leader is the Professor Jesús M. Soria of the Alicante University.

Jesús Soria (in the 90’s) pointing to a palm-tree-gypsum near Benejúzar (Alicante).
Team Members:
Antonio Estévez
Alfonso Yébenes
José Antonio Pina
José Enrique Tent-Manclús
Ignacio Fierro
José Francisco Baeza-Carratalá
Alice Giannetti
Hugo A. Corbi
Juan J. Peral

![]()
Alicante Messinian Group
This blog is the continuation of the web page of the Alicante Messinian Group.
www.messinianalicante.com

Recent Comments