Students of the third course of Geology of the Alicante University within the subject of Regional Geology: visit Jarafuel triassic section, Montealegre del Castillo triassic section, Sot de Chera Jurassic section and Monduver geologic dome.
The picture shows the in the Chulilla health resort section near by Sot de Chera (Valencia province)

University of Alicante geology students near Sot de Chera.
Tags: Geology, University Alicante
This year the students of the third course of Marine sciences of the Alicante University visit the Sorbas basin, Cabo de Gata, and Mesa de Roldán on November 4th and 5th of 2021.

University of Alicante Marine Geology course students (2021-22)
The picture show the student visiting the Torregarcia beach and the Roman remains of a cetaria (a marine factory).
The professors in charge were Antonio Estévez, Manuel Martín-Martín and José Enrique Tent-Manclús.
The Monduver massif located 50 km to the south-east of Valencia city has a well-developed Karst geomorphology and where it stands out the Barx Polje. The Highest point of the Massif, Moduver 841 m a.s.l., is just 10 km inland from the Mediterranean coast. The Parpallo Cave located at the southern part of the polje is one of the most important Palaeolithic sites, not only in Spain, but in the world due to the collection of decorated plaquettes of Palaeolithic portable art. The collection covers the entire archaeological sequence from the Gravettian to the Magdalenian periods (26000–11000 years BP). The Barx polje has two subbasins: Barx to the north and La Drova to the south, and the second one is partly capture at the south by the Caldereta Creek. In this work is proposed than the La Drova subbasin was formed by a major landslide of the western side of the Monduver massif producing a natural dam of the Les Voltes Creek. Lately, the La Drova subbasin was filled by the terra rossa product of the limestone dissolution of the polje basin, as can be see today in the sides of the captured La Drova subbasin. The filling of the subbasin was to a level higher that the Barx subbasin as the today drainage of the uncaptured La Drova runs to this last one. Three 3d model landscape figures has been made to explain its evolution. The archeological sequence of the Parpallo Cave could be related with the infilling of the subbasin period and the landslide could take place before the human use of the cave.

Panoramic view from the Parpalló Cove entrance. The face-shape mountain is the paleoslide
Cite as: Tent-Manclús, J. E. y Jordá Guijarro, J. (2021): Monduver Paleolandslide as its relationship with the human use of the Parpalló Cave (Valencia province, Spain). LAC1+1, Landscape archaeology conference, 8-11 june 2021 Book of Abstracts. 14 p.
Tags: Barx Polje, Geomorphology, Holocene, Monduver massif, Paleolandslides, Parpallo cave
The Alcudia archaeological site correspond to the ancient city of Ilici (Roman Colonia Iulia Ilici Augusta). The site consists of an succession of Iberian, roman, byzantine, Visigoth and Moorish settlements (since the 5th centuries BC to the 8th century AC) on the top of a hill that in the 1st century BC was surrounded by a wall and nearby the Vinalopó river mouth in the Bajo Segura Bay. This bay was known in Roman times as the sinus Ilicitanus. Different clays of the area nearby to the site have been sampled, analysed and then compared with analysis of some pottery remains of the different periods of the Alcudia settlement. The clays have been analysed by micro-X Ray fluorescence (µXRF) and by infrared spectrometry by Fourier transformed (ATR-FTIR), as crude, mixed in different proportions, and then heated to a different temperature until 900ºC. The old remains show affinities with the clays sampled from the Eastern part indicating a W-E influence, as the northern coast of the sinus ilicitanus. During the roman period and later the pottery was made using the nearby clays located just to the north of the site clearly influenced by the via Augusta. The topography to the north of the site is formed by low but steep hills and small creeks that possibly limit the commerce to the north, so the commerce follows the marine plain coast of the northern sinus ilicitanus. The opening of the roman road via August along the Vinalopó Valley favoured the use of pottery clays from the interior of the territory.
Paleogeographic map of the area north of the sinus ilicitanus. The clays used for pottery were obtained from the area parallel to the north shore of the sinus. Pottery: Variable carbonate no sílices
Cite as: Jordá Guijarro, J., Cerdán Sala, M., Sánchez-Sánchez, A., Ronda Femenia A., Tendero Porras, M. and Tent-Manclús, J. E. (2021): Pottery clays quarries in the domain territory of ancient Ilici (Alicante, SE Spain). LAC1+1, Landscape archaeology conference, 8-11 june 2021 Book of Abstracts. 45 p.
Tags: Elche, Holocene, illice, pottery clay, sinus ilicitanus, via augusta
Change on the coastline of the Southern Alicante region
Tent-Manclús, J. E.
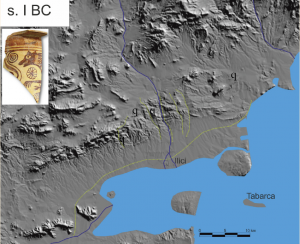
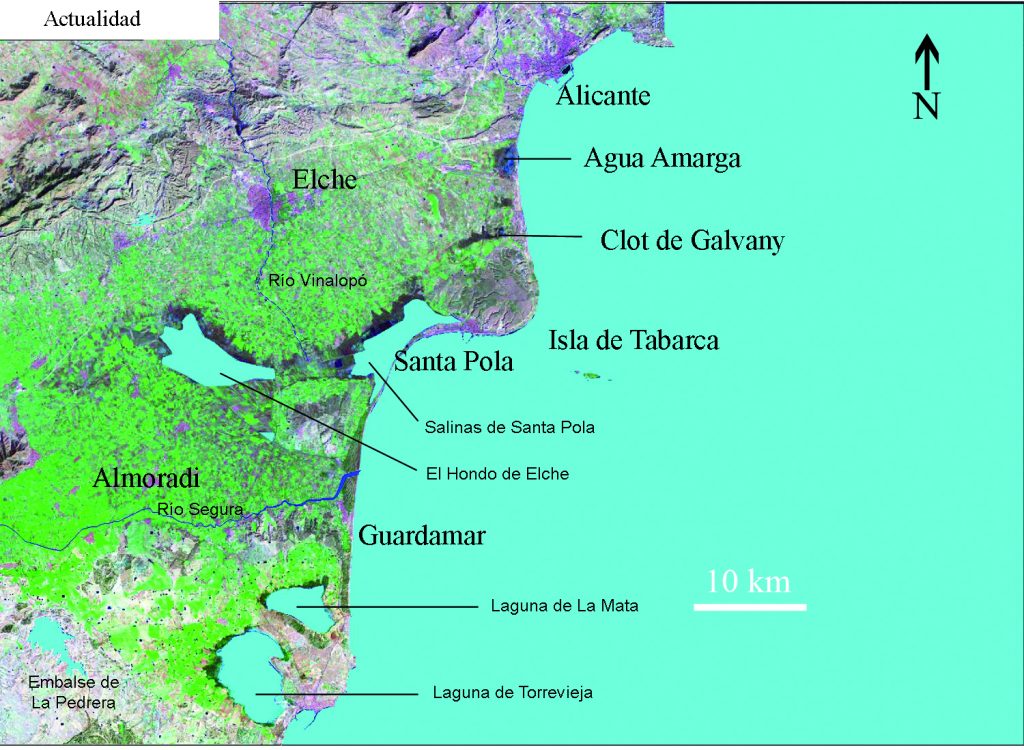
The Bajo Segura Region is a plain dominated by the Segura River with the mouth towards the Mediterranean Sea on Guardamar. The Hondón and Salinas de Santa Pola lagoons inside the plain, are the remains of a major marine bay known in the Roman times as the sinus ilicitanus. Offshore the Region geophysical data has permitted to model the coastline change and study the underwater marine terraces. 183 maps have been made from the 15000 BP until Today (100 year apart the older ones and 25 years for the recent ones). The coastline has migrated between 20 and 50 km onshore and then offshore about tens of meters to 35 km shaping today coastline. The sea level at 15000 BP was located 70 m below today sea-level (btsl). During the Younger Dryas (13500 to 11700 BP) varies around 60-66 m btsl then increase until 21 m btsl and stopped during the 8.2 ka event (8500 BP to 8200 BP). Again, the level increases first fast and then slowly but moving fast horizontally (near 35 km for 1000 years) developing the sinus ilicitanus when the marine transgression enters the plain about 4200 BC. The Segura River developed a delta in the today area of Orihuela, 35 km to the East of its today river mouth in Guardamar. The river until around the year 1400 AD developed a fingering (bird-foot) quite variable delta in the center of the sinus, most of the major irrigation canals (acequias) could correspond to the ancient delta-channels. Around the XV century major events changed the sinus basin producing the river mouth move to the present location. The sinus open communication with the Mediterranean Sea was closed around the XVIII century by sand bars, first the southern of Guardamar and then the northern on Santa Pola.
Cite as:
Tags: Alicante coast, coasline change, Holocene, sea-level change, Segura River, sinus ilicitanus
This year the students of the third course of Marine sciences of the Alicante University, could not go for a field trip on November as previous years, visiting the Sorbas basin, Cabo de Gata, and Hoyazo de Nijar. When the courses ended on May most of the students did the visit. The visit took place on 27 and 28 of May of 2021.
The field trip was strange because of the new normality and also the rain comes to Almeria during the visit.

Alicante University students of Marine Sciences in the Sorbas basin
The professors in charge were Juana Jordá, and José Enrique Tent-Manclús.
Tags: Holocene
Students of the third course of Geology of the Alicante University within the subject of Regional Geology: visit La Albufera de Valencia. First we have a view of the Ermita dels Sants de la Pedra (Sueca) and then the Silla Port. The evolution of the Albufera from the Middle Ages when was connected with the sea to the today lake was explained. The Moorish expulsion using the today disappeared Sueca Port was pointed as the one of the subject to work with.

Students watching the smoke produced by the rice fields fires
Tags: Albufera, Alicante University, Holocene
The change of the coastline in the southern part of the province of Alicante for the last 15,000 years have been modeled and described in the work of Tent-Manclus (2013). It is obtained by integrating data from different sources and especially high-resolution seismic profiles of the nearby marine continental shelf. Ten periods have been distinguished in the evolution of the Bajo Segura coastline : 1) 15000 BP- 14600 BP: rapid rise; 2) 14600 BP to 13500 BP: the Older Dryas, slow rise; 3) 13500 BP to 12700 BP: stability and descent; 4) 12700 BP to 11700 BP: the Younger Dryas, descent of the sea level; 5) 11700 BP to 11300 BP: very fast rise; 6) 11300 BP a 8500 BP: rapid rise; 7) 8500 BP = 6500 AC to 8200 BP = 6200 AC: 8.2 ka event stabilization; 8) 6,200 BC to 4,000 BC: rise, Tabarca Island is form; 9) 4000 BC to 3000 BC: the slow rise due to subsidence sinus ilicitanus forms and 10) 3000 BC to the present day 2000 AD: the sinus ilicitanus desiccates.
Map of the southern Alicante coast area used in the work
Maps at different times are show selected from the 164 maps that have been made. This is a first version of how the coastline evolves in the region. This work It can be a starting point to improve knowledge of the historical landscape evolution..
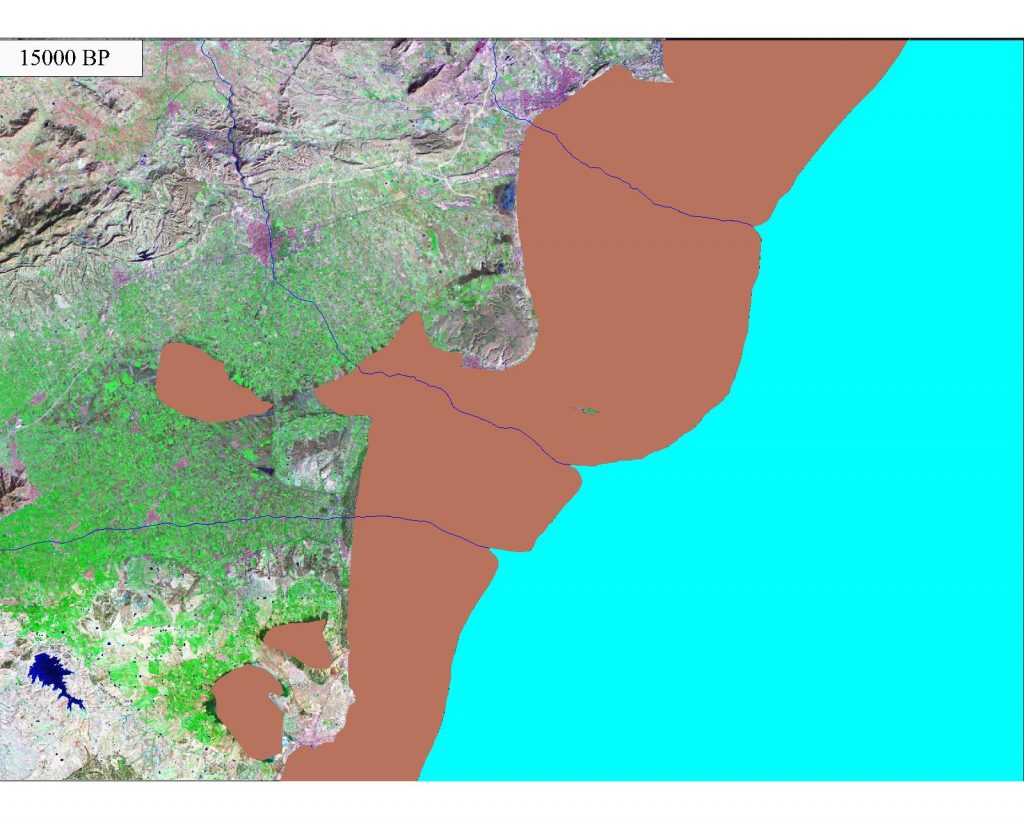
Map of the South Alicante coast 15000 BP. The sea level was 70 m below today level. The rivers Vinalopo and Segura were running parallel.
The sinus ilicitanus dries until today, the rest of which are the lagoons of the Fondo and Santa Polo Salinas.
Cite as: Tent-Manclús, J. E. (2013): Cambios de la línea de costa en el Bajo Segura (S de Alicante) en los últimos 15.000 años. Estudios geográficos, 74 (275): 684-702. Doi: 10.3989/estgeogr.201324
Tags: Alicante coast, Bajo Segura, coastal maps, Holocene, sea-level change
Last February 6, 2020, students in the third year of the Marine Geology Degree carried out practices aboard the “Guadalupe” boat. This 12m-length boat assigned to the City of Dénia by the Court of Instruction number 3 of Dénia by the Judge Javier Reyes, is dedicated not only to surveillance tasks of the Marine Reserve of the Cape of San Antonio, but also to operations of pollution control and scientific monitoring of the protected area.
The practice was focused on different aspects of “Geophysics and geophysical prospecting”, under the supervision of Professor José Enrique Tent Manclús. Within a research program of the Multidisciplinary Institute for the Study of the Environment “Ramón Margalef” (IMEM) of the University of Alicante that together with the City Council of Dénia and the National Parks Autonomous Organization operate the Montgó-Dénia Scientific Station (ESCIMO-Dénia).
Taking advantage the good conditions at sea, marine geophysics equipment was tested, specifically a high-resolution marine boomer equipment linked to a differential GPS. The practice was to leave the port, perform the boot sequence and navigate acquiring data. Through sound waves, this equipment allows interpreting the geometry of the layers below the seabed using the same principle as the medical ultrasound, which was precisely developed from this marine technology. Like 2d ultrasound, experience and practice are required for interpretation.
In this first marine campaign, it has been tested the equipment adjustment settings to obtain a better results. The students have been able to see in situ how the equipment studied in the classroom works and, above all, have a first contact with work in the marine environment, much more hostile than the terrestrial to carry out any study.

Student inside the Guadalupe boat during the sailing.
The marine area of the San Antonio Cape is the extension of the rocky massif of Montgó. The equipment is intended to see the thicknesses of the sandbars and / or sediments trapped by the Posidonia oceanica, above the rocky bottoms. In a future campaign will be locate and delimit the bodies of sand and how they change to areas without sand. In the deepest parts it is expected to be able to locate some level of marine terrace formed when the sea level was lower as a result of the last glaciation. At the last glacial maximum, 18,000 years ago, the sea level was 120 m below the current level. A warming began that caused a rise of 1 to 2 cm a year, but about 14,000 and 11,700 years ago the global climate cooled, which stopped the ascent. This generated rocky coasts that are currently marine terraces between 70 and 60 m deep. After this cold interval known as the “Younger Dryas”, the level rose to reach its current position about 6000 years ago. Younger Dryas terraces have already been located in the coastal area Tabarca Island (South coast Alicante) and in the near future it is intended to study these terraces in the area of San Antonio Cabe.
This study of marine geology together with others of a biological and physical-chemical nature, promoted by the Montgó Scientific Station (ESCIMO-Dénia), not only seeks to improve the knowledge of a very interesting area at the geological level but also to lay the foundations for a proposal to expand and improve the fishing resources for the artisanal fleet around the Marine Reserve as well as the promotion of sustainable activities such as diving.
Tags: Denia, ESCIMO-Denia, Holocene, IMEM, University Alicante
As the last year, students of the third course of Marine sciences of the Alicante University visit the Sorbas basin, Cabo de Gata, Hoyazo de Nijar, and Cerro del Espíritu Santo of Vera. The visit took place on 7 and 8 of November of 2019.
The visit was organized by Antonio Estévez, Santiago Moliner, and José Enrique Tent-Manclús.

Student group before entering the Sorbas Cave.
Recent Comments