The project will study changes in the coast-line, coastal risk and coastal aquifers, an its logo is:
![]()
The leadership of the project is carried out by researches of the MIES is a Multidisciplinary Research Institute for Environmental Studies “Ramón Margalef” (Instituto Multidisciplinario para el estudio del Medio “Ramon Margalef”, IMEM in Spanish) of the Alicante University and integrate researches of 10 universities and research institutions.
![]()
The EU funded ThinkInAzul programme supported by MCIN (Ministerio de Ciencia e Innovación) with funding from European Union NextGenerationEU (PRTR-C17.I1) and by Generalitat Valenciana
![]()
![]()
Rogério Portantiolo Manzolli (Universidad Autónoma Madrid), Claudi Blasco (Universidad Alicante), José Enrique Tent-Manclús (Universidad Alicante) and Carlos Arteaga Cardenau Universidad Autónoma Madrid) in Guardamar making a manual borehole.
Tags: MOLICO-CV
Last March 4th, 2023, students in the third year of the Geology Degree carried out practices aboard the “Rosa de Abril” boat from the Torrevieja harbor . The practice was focused on different aspects of “Geophysics and geophysical prospecting”, under the supervision of Professor José Enrique Tent Manclús. This academic we have returned to sail as before the pandemic.

Tags: Geology, Geophysics, Torrevieja, University Alicante
At 30 nautical miles to the E of Cabo de Santa Pola (about 55 km) there is a submarine seamount called by the southern Alicante fishermen as the area of the Patibulo (the Gallows). It is an area with a rocky and elevated seabed, with abundant fishing.
Curiously, not only are fish abundant, but from time to time the seismographs of the Alicante University Seismic Record Unit record earthquakes there and along the imaginary line that joins it with Cabo de Santa Pola, to the north of Tabarca Island. In the coastal area to the north of said island, an active fault has already been located through high-resolution seismic as a possible cause of these. It was missing to know if in the Patibulo, said fault continues or another one also active.
Throughout the year 2022, attempts have been made to reach the area from Torrevieja with marine seismic equipment to study the area. It started in the month of April and something always happened, a fuse, a cable, a blown chip, bad weather… After much insistence and returning empty-handed, on December 6, 7 and 8, there was a good weather window and the appliances worked. From the port of Torrevieja with the boat “Rosa de Abril”, we were making some data collection lines in the direction of the Patibulo and on the last day we reached our objective. Almost 80 km of data lines were obtained over 3 days.
El Patíbulo is the westernmost seamount of an alignment of elevations called Alicante Highs, after the geological map of the Alicante platform (Catafau Alcántara et al., 1994). There are at least three needles whose base is 300 m deep and which rise to about 135 m, the most to the W (Patíbulo). The second is the highest and reaches 125 m (some fishermen say 90 m), the third at 154 m. The image of the geological map of the Alicante platform is attached. To the north of these thresholds begins a submarine canyon that is “unofficially” known as the “Alicante canyon” and that descends to the plain to the north of these.
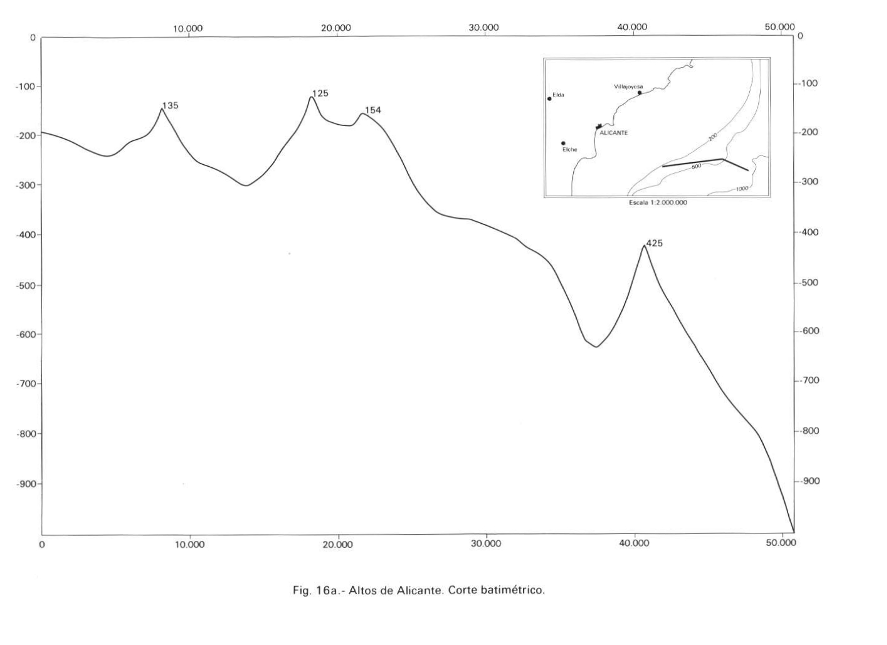
Alicante Heights, this profile does not pass through the tops of the heights, it is a bathymetric profile where some of them are located (Taken from Catafau Alcántara et al., 1994).
The high-resolution seismic image shows a fault scarp on the seafloor, indicating that the fault is currently active. The darker area marked by the plane is a more cemented area, probably due to the flow of fluids along the plane. Layered reflectors are also observed, indicating that they are sedimentary rocks and not volcanic rocks like other seamounts in the southern Balearic Islands.

Fault in the Gallows area. A fault scarp is observed on the surface of the seabed and its prolongation in depth. The layers on the right cut off at the fault. Raw image obtained during acquisition.
With the preliminary data that has been obtained, it can be said that the Patibulo is a structural height raised by faults. They are active because they protrude from the bottom and because it seems that they are associated with leaks of fluids that mineralize near the fault plane.
We look forward to studying the interesting data obtained.
Reference: Catafau Alcántara, E., Gaytán de Ayala y Bolín, M., Pereda Alonso, I., Vázquez Garrido, J.T., Wandossell Santamaría, J. (1994): Mapa geológico de la Plataforma Continental Española y Zonas adyacentes. Escala 1:200 000. Hoja nº 72-73(Elche-Alicante). ITGE, 73 p.
Tags: Alicante coast, Alicante Highs, Patibulo, Torrevieja
This academic year 2022-23 we have returned to Teruel Paleozoic as before the pandemic. The geology students of Alicante University third course within the subject of Regional Geology: visit Montealegre del Castillo triassic section (Albacete), Aguilon Jurassic section (Teruel), Murero Cambrian Rambla Valdemiedes section (Zaragoza), Luesma Ordovicic to Triassic section (Teruel), Santa Cruz de Nogueras museum “Paleozoic Seas Museum” (Teruel), Ojos Negros iron-ore quarry (Teruel) and Checa-Orea road Ordovicic-Silurian section (Guadalajara).
The picture shows all the students on the Tertiary erosional surface on top of the Murero section (Zaragoza province).

Alicante University Student in Murero top section. Panoramic view of the Iberian Cordillera.
The second picture show students in the Murero Cambrian Rambla de Valdemiedes section.

Alicante University geology third course students in the Rambla de Valdemiedes, cambrian of Murero.
This year the students of the third course of Marine sciences of the Alicante University visit the Sorbas basin, Cabo de Gata, and Mesa de Roldán on November 3th and 4th of 2022.

University of Alicante Marine Geology course students (2022-23).
The picture show the student visiting the Faro de la Mesa de Roldan (the lighthouse).
The professors of the subject were Antonio Estévez, Manuel Martín-Martín and José Enrique Tent-Manclús.
Tags: Almeria, Betic cordillera, marine sciences
Last september 16th 2022 in the IMEDMAR facilities of the Universitat Católica de Valencia took place the first meeting of the WP1 of the Thinkinazul project.
The meeting did not start quite well because 3 days before the meeting a fire destroyed the entrance of the building so it had to be moved to the Calpe Nautic Club, Belliure brothers meeting room. It was our first meeting, our kicking point trying to find synergies with the different subgroups.

After a complete study of the sediment samples from Ocean Drilling Program (ODP) Sites 999 and 1237 in the Caribbean Sea and Eastern Pacific Ocean in order to monitor the evolution of ceratoliths. The First Occurrence of Amaurolithus delicatus marks the bottom of the Messinian Stage. The first ceratholith, A. primus, has two arms and a horseshoe shape with marked laths, and is stable and concave upwards having three blades (sinistral=blue, median=red, and dextral= green).
Early robust A. primus evolved into stylised forms and then to Amaurolithus delicatus (7.226 Ma), an almost plain horseshoe ceratolith with two arms. The left arm, usually the longer one, comes from the sinistral wing of A. primus and has a characteristic flattened omega section (Ω) without laths. There is a distribution overlap between A. primus and A. delicatus; the highest occurrence of the first one, at 6.282 Ma, is a newly proposed bioevent for the Messinian. After this, Amaurolithus does not have laths on its longer left arm and should be included in A. delicatus.
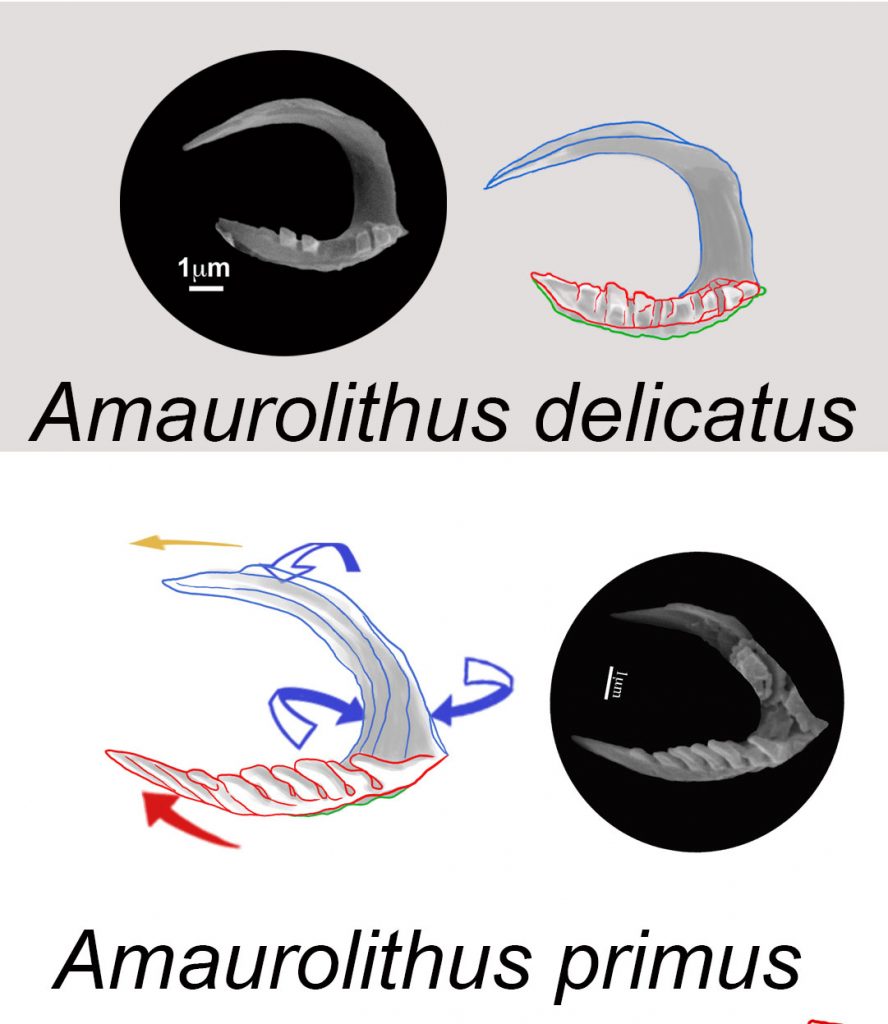
Cite as: Lancis, C., Tent-Manclús, J.-E., Flores, J.-A., 2022. Origin and evolutionary trends of the Neogene genera Amaurolithus and Nicklithus (calcareous nannofossils). Mar. Micropaleontol. 175, 102156. https://doi.org/https://doi.org/10.1016/j.marmicro.2022.102156
Tags: A. delicatus, A. primus, Amaurolithus, nannplankton evolution
Sediment samples from Ocean Drilling Program (ODP) Sites 999 and 1237 in the Caribbean Sea and Eastern Pacific Ocean were studied to monitor the evolution of ceratoliths from 7.4 Ma to 6 Ma. Orthorhabdus rugosus shows high variability at the end of the Tortonian to the Early Messinian (7.35 Ma to 6.91 Ma), resulting in the Amaurolithus (7.354 Ma) and Nicklithus branches (6.985 Ma). Orthorhabdus rugosus is an ortholith with three blades (sinistral, median, and dextral). The first ceratholith, A. primus, has two arms and a horseshoe shape with marked laths, and is stable and concave upwards. Its sinistral arm is formed from the sinistral blade of Orthorhabdus rugosus and the right arm is formed from the other blades.
The sudden appearance of A. primus in the sedimentary record at around 7.35 Ma was observed in both studied sections . The Figure shows the set modifications of O. rugosus producing the early A. primus. The modifications are not a gradual evolutionary sequence of intermediate progressively modified forms but occur simultaneously as they are present from the beginning of the first recorded specimens. Amaurolithus primus coexisted with the O. rugosus ancestor in the studied samples.
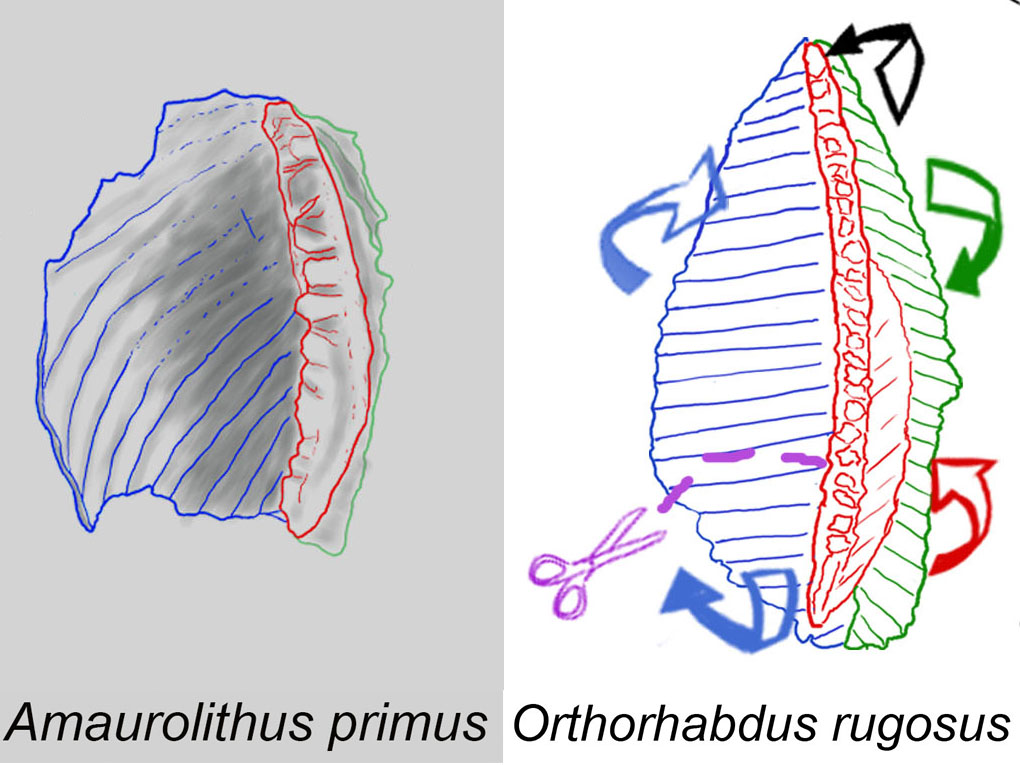
Modification on O. rugosus: Shortening of the specimen; Sinistral blade (blue): Lateral lengthening and its end portion curved upwards, concave to the observer; Dextral blade (green): Rotates backward; Median blade (red): Elevates and becomes more robust showing more marked teeth.
Cite as: Lancis, C., Tent-Manclús, J.-E., Flores, J.-A., 2022. Origin and evolutionary trends of the Neogene genera Amaurolithus and Nicklithus (calcareous nannofossils). Mar. Micropaleontol. 175, 102156. https://doi.org/https://doi.org/10.1016/j.marmicro.2022.102156
Tags: Amaurolithus, Orthorhabdus
Messinian Group will study the recent marine sediment on the Alicante coast. The new EU funded Thinkinazul project of the Generalitat Valenciana, coded as: GVA/Thinkinazul/2021/039.
The project will study changes in the coast-line, coastal risk and coastal aquifers. It has started on January 1, 2022 and will be active until November 18, 2024. The advances of the research will be announced in future posts of this blog. The leadership of the project is carried out by researches of the MIES is a Multidisciplinary Research Institute for Environmental Studies “Ramón Margalef” (Instituto Multidisciplinario para el estudio del Medio “Ramon Margalef”, IMEM in Spanish) of the Alicante University and integrate researches of 10 universities and research institutions.
Generalitat Valenciana Thinkinazul Project “aquaCHANGE” have been divided in 7 Working Party (WP) and framed in WP1: “Observation and monitoring of the marine and coastal environment”.

Tags: Alicante coast, Alicante University, Holocene, IMEM
Students of the third course of Geology of the Alicante University within the subject of Regional Geology: visit Jarafuel triassic section, Montealegre del Castillo triassic section, Sot de Chera Jurassic section and Monduver geologic dome.
The picture shows the in the Chulilla health resort section near by Sot de Chera (Valencia province)

University of Alicante geology students near Sot de Chera.
Tags: Geology, University Alicante
Recent Comments